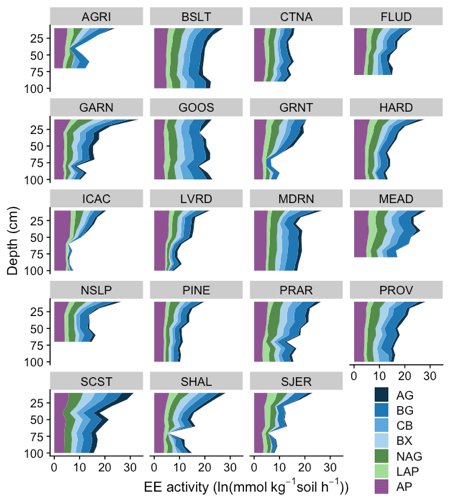
Stabilization of microbial-derived products such as extracellular enzymes (EE) has gained attention as a possibly important mechanism leading to the persistence of soil organic carbon (SOC). While the controls on EE activities and their stabilization in the surface soil are reasonably well-understood, how these activities change with soil depth and possibly diverge from those at the soil surface due to distinct physical, chemical, and biotic conditions remains unclear. We assessed EE activity to a depth of 1 m (10 cm increments) in 19 soil profiles across the Critical Zone Observatory Network, which represents a wide range of climates, soil orders, and vegetation types. Activities of four carbon (C)-acquiring enzymes (α-glucosidase, β-glucosidase, β-xylosidase, and cellobiohydrolase), two nitrogen (N)-acquiring enzymes (N-acetylglucosaminidase and leucine aminopeptidase), and one phosphorus (P)-acquiring enzyme (acid phosphatase) were measured fluorometrically along with SOC, total N, Olsen P, pH, clay concentration, and phospholipid fatty acids, which we used to characterize the microbial community composition and biomass (MB). For all EEs, activities per gram soil correlated positively with MB and SOC; all of which decreased logarithmically with depth (p < 0.05). Across all sites, over half of the potential soil EE activities per gram soil consistently occurred below 20 cm for all measured EEs. Activities per unit MB or SOC were substantially higher at depth (soils below 20 cm accounted for 80% of whole-profile EE activity), suggesting an accumulation of stabilized (i.e., mineral sorbed) EEs in subsoil horizons. The pronounced enzyme stabilization in subsurface horizons was corroborated by mixed-effects models that showed a significant, positive relationship between clay concentration and MB-normalized EE activities in the subsoil. Furthermore, the negative relationships between soil C, N, and P and C-, N-, and P-acquiring EEs found in the surface soil decoupled at 20 cm, which could have also been caused by EE stabilization. This suggesting that EEs do not reflect soil nutrient availabilities at depth. Taken together, our results suggest that deeper soil horizons hold a significant reservoir of EEs, and that the controls of subsoil EEs differ from their surface soil counterparts.